Il sistema nervoso è la sede dell'assunzione, dell'elaborazione e trasmissione delle informazioni; in altre parole è un sistema di regolazione delle funzioni coporee e "fa concorrenza" al sistema endocrino. Il sistema nervoso è un complesso sistema elettrochimico, comnposto da oltre 1000 miliardi di cellule, che viaggia a 400 km all’ora e che assorbe il 20% dell’ossigeno del nostro organismo.
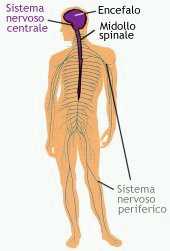
Encefalo e midollo spinale formano il sistema nervoso centrale o asse cerebrospinale che è in grado di raccogliere, trasmettere e integrare le informazioni. Il sistema nervoso periferico, formato dai nervi spinali, dai nervi cranici e dal sistema vegetativo, svolge essenzialmente la funzione di trasmissione del segnale: segnali afferenti da un'unità periferica (organo) o efferenti verso un'unità periferica decorrono in fibre separate (assoni) generalmente raggruppate in un fascio di conduzione unitario (nervo). Un nervo contiene esclusivamente assoni, cellule di Schwann e tessuto connettivo. I corpi delle cellule nervose sono raggruppati nei gangli nervosi del sistema nervoso periferico e nei nuclei del midollo spinale e dell'encefalo, che costituiscono dei pool neuronali con specifiche caratteristiche di organizzazione ed elaborazione dei segnali.
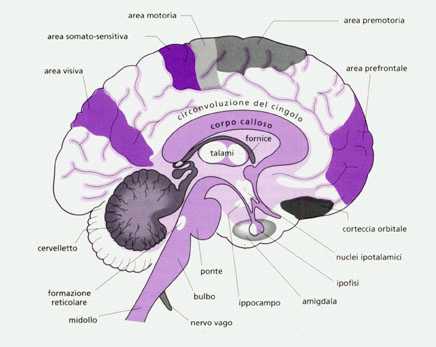
L'encefalo, con un peso di 1,3-1,5 kg, è il secondo organo più pesante del corpo dopo il fegato. A riposo, il 25% dell'energia metabolica viene utilizzata per rifornire il cervello (8-10% in più rispetto agli altri primati non umani). Le strutture più antiche ("inferiori") si trovano in profondità (inferiormente e immediatamente sopra il forame occipitale), mentre le componenti più recenti ("superiori") si collocano sopra di esse.
Il midollo allungato (bulbo) è la parte più antica e la sua struttura si continua col midollo spinale (di cui ricorda la struttura metamenrica). Nel midollo allungato i fasci motori, provenienti dalla corteccia cerebrale e diretti al midollo spinale, formano le piramidi, dove avviene la loro decussazione (incrocio a X).
Il bulbo o midollo allungato, attraverso confini ben delineati, continua col ponte (ponte di Varolio) a cui si unisce il mesencefalo (o mesoencefalo), che assume una posizione mediana fra le regioni cerebrali antiche e recenti. All'interno del mesencefalo è presente il nucleo rosso. Il midollo allungato e il ponte formano il romboencefalo. Il romboencefalo più il mesoencefalo fomano il tronco encefalico.
I nervi cranici (nervi encefalici) sono un gruppo di nervi che invece di avere origine dal midollo spinale, partono direttamente dal tronco encefalico costituendo l'innervazione motrice e sensitiva della testa nonchè l'innervazione viscerale, sia in uscita che in entrata (efferente e afferente), degli importanti centri del sistema nervoso parasimpatico che controllano l'attività dei visceri della testa, del collo, del torace e dell'addome. Nell'anatomia umana ci sono dodici paia di nervi cranici (destri e sinistri), numerati dall'alto verso il basso con numeri romani. Essi hanno un'organizzazione più complessa rispetto ai nervi spinali. Anche i nuclei dei nervi cranici (nuclei encefalici), ossia le zone in cui sono situati i corpi cellulari dei neuroni corrispondenti, sono più complessi di quelli spinali. La sostanza grigia, a differenza del midollo spinale, non è raccolta in un'unica formazione ma è suddivisa in vari nuclei in parte ai nervi encefalici. Quasi tutti i nervi cranici sono connessi a dei nuclei di materia grigia all'interno del tronco encefalico e gli assoni da e per i nervi cranici incontrano le loro sinapsi all'interno di questi nuclei. Come nel midollo spinale, anche i nuclei del tronco encefalico sono divisi in anteriori motori (somatici) e posteriori sensoriali. Nel tronco encefalico vi sono inoltre nuclei non direttamente connessi all'origine dei nervi encefalici, detti nuclei propri, e la formazione (sostanza) reticolare (antica formazione nervosa dalla citoarchitettonica caratteristica, costituita da varie tipologie neuronali aggregate in piccoli nuclei, nell'ambito di un complesso reticolato di fibre nervose che si estende per tutto il tronco encefalico). Fra quesi nuclei citiamo, per l'importanza rivestita nel sistema dell'equilibrio e postura, i nuclei vestibolari e i nuclei oculomotori.
Il ponte presenta importanti connessioni col cervelletto che è la seconda regione dell'encefalo più voluminosa dopo il "cervello superiore". Esso si appoggia posteriormente al romboencefalo piegandone anteriormente l'asse, così che il ponte si trova ad appoggiarsi direttamente sulla base cranica. Il cervelletto è diviso in due emisferi cerebellari divisi dal verme, sulla cui faccia anteiore è presente il flocculo. Esso è costituito da regioni antiche (verme, flocculo e il quarto anteriore dei due emisferi) e regioni recenti (compartimenti posteriori degli emisferi), che rappresentano la parte più estesa, con funzioni diverse. All'interno della sostanza bianca, in profondità, sono situati i nuclei cerebellari (ammassi di sostanza grigia) di cui, il nucleo dentato, risulta particolarmente sviluppato. La sostanza grigia della corteccia cerebellare (ca. 1 mm di spessore) si ripiega in superficie per accogliere il maggior numero possibile di neuroni.
Superiormente al mesoencefalo (mesencefalo) troviano il diencefalo composto fondamentalmente dal talamo destro e sinistro e dall'ipotalamo posto al centro, quest'ultimo è collegato con la ghiandola endocrina ipofisi (sistema neuroendocrino), con la sostanza reticolare e col sistema limbico. Il sistema o lobo limbico rappresenta quei giri corticali, filogeneticamente più antichi, che circondano ad anello il tronco encefalico e comprende l'amigdala, l'ippocampo, i corpi mammillari ed il giro del cingolo o cingolato (immagine). Il sistema limbico è un circuito costituito da un insieme di centri corticali e subcorticali fra loro interconnessi da proiezioni di fasci di fibre nervose (tra cui le più importanti sono la fornice che unisce ippocampo e diencefalo e la stria terminale che collega l'amigdala all'ippocampo) che trasportano specifici neurotrasmettitori. Notevole è l'importanza dell'insieme ipotalamo-sistema limbico relativamente a numerose funzioni vitali dell'organismo.
Il telecenfalo, considerato dai fisiologici e dai neuroclinici il "cervello" in senso stretto, è la parte di gran lungo più voluminosa dell'encefalo e ricopre tutte le precedenti strutture. Il telencefalo risulta diviso in due emisferi (destro e sinistro) uniti tra loro profondamente, all'interno della scissura interemisferica (in cui penetrano le meningi), da fasci di fibre nervose che formano un nucleo di sostanza bianca chiamato corpo calloso. Ogni emisfero è diviso in 4 lobi (frontale, parietale, temporale e occipitale) in cui si identificano aree (aree di proiezione) con funzioni motorie o sensitive prevalentemente specifiche.
La parete degli emisferi cerebrali è la corteccia cerebrale costituita da sostanza grigia ricca di neuroni che riveste la sostanza bianca formata da fibre nervose (assoni). Per far fronte all'aumento del numero delle cellule cerebrali nel corso dello sviluppo evolutivo (nell'uomo sono ca. 10 miliardi), la corteccia cerebrale, come nel cervelletto, ha dovuto ripiegarsi su se stesso, formando le circonvoluzioni (nel regno animale è possibile dimostrare che all'aumentare delle capacità intellettive corrisponde un aumento della complessità delle circonvoluzioni cerebrali).
L'organizzazione dell'encefalo non si differenzia in modo sostanziale da quella del midollo spinale ma solo nel telencefalo e nel cervelletto è la sostanza grigia (corpi di cellule nervose e fibre nervose amieliniche) a circondare la bianca (fibre nervose mieliniche). Il vantaggio di questo tipo di organizzazione, rispetto a quallo presente nel midollo spinale e nel troncoencefalo (sostanza bianca all'esperno e grigia all'interno), è di consentire vie di collegamenti più brevi (questo vantaggio è sfruttato anche nei computer moderni in cui le chips sono poste esternamente e il cablaggio internamente). Analogamente al cervelletto, all'interno della sostanza bianca, in profondità, sono situati i nuclei della base (globo pallido, sostanza nera, putamen e nucleo caudato; quest'ultimi due spesso compresi nel "corpo striato").
Quanto più una porzione cerebrale è recente, tanto più complesse sono le sue funzioni. Il midollo allungato contiene i centri vegetativi vitali (in particolare repiratori e circolatori) e collabora strettamente con il ponte, il cervelletto e il mesencefalo in quella che sembra essere la loro funzione principale ovvero la regolazione dell postura in statica e in movimento (situazione simile è presente nei vertebrati inferiori). Il talamo è il centro riflesso delle emozioni che provocano il pianto e il riso controllandone la motilità relativa e la sensibilità degli imput ricevuti dai recettori periferici,
L'ipotalamo e il sistema limbico, con il quale è funzionalmente e strutturalmente collegato, presiedono a quei meccanismi vitali che hanno lo scopo di mantenere costanti le condizioni dell'ambiente interno (omeostasi) e di provvedere alla conservazione dell'individuo e della specie, esercitando il controllo sulle sue emozioni e sulla sessualità: regolazione del sistema nervoso autonomo e dell'apparato endocrino, della temperatura corporea, del ciclo sonno/veglia, della frequenza cardiaca, della pressiona arteriosa, dell'osmolarità del sangue, dell'assunzione di cibo e acqua, della secrezione, acida dello stomaco, del metabolismo dei glicidi e dei grassi, delle emozioni e delle funzioni sessuali. L'ippocampo e' inoltrre importante per l'apprendimento e la memoria. La caratteristica peculiare dell'ipotalamo è quella di ricevere degli imput direttamente dall'esterno del cervello. L'ipotalamo infatti riceve direttamente il segnale luce-buio (tratto retino-ipotalamico) e quello olfattivo (via olfattiva basale), in più, esso costituisce, tramite un fascio di fibre nervose e una rete di vasi sanguinei (eminenza mediana dell'ipotalamo) che oltrepassano la barriera emato-encefalica (barriera, costituita dai capillari cerebrali, dai plessi corioidei e dall’aracnoide, che protegge il cervello dal passaggio di batteri, virus e gran parte della molecole circolanti nel sangue), la parte posteriore della ghiandola endocrina ipofisi, detta neuroipofisi. La neuroipofisi non risulta quindi essere in realtà una ghiandola ma un'estroflessione cerebrale formata cioè da tessuto nervoso. Questo collegamento diretto sistema nervoso centrale-apparato endocrino (sistema neuroendocrino) assume grande rilievo nella regolazione generale dei processi vitali. L'ipotalamo inoltre è esso stesso anche una ghiandola endocrina i cui ormoni, agenti sull'adenoiposi (fattori ipotalamici), creano con essa un doppio legame: strutturale (neuroipofisi) e umorale, tramite ormoni (adenoipofisi). L'ipotalamo grazie alle sue connessioni con la neocorteccia, col sistema limbico, con la sostanza reticolare, col sistema nervoso vegetativo e col sistema endocrino è cosiderato da molti studiosi la struttura limite tra somatico e psichico, quella cioè in grado di commutare il segnale pschico in chimico e viceversa. E' grazie all'ipotalamo che gli aspetti mentali, emotivi e istintivi trovano espressione nel soma.
Nel telencefalo, infine, hanno sede le funzioni cerebrali "superiori" quali il pensiero, il linguaggio, la programmazione motoria ecc.
Il canale formato dalle vertebre costituisce un'ottima protezione per il delicato midollo spinale. Il midollo spinale ha forma cilidrica, con diamentro medio 8-10 mm, e si estende dal grande forame occipitale (base cranio), continuandosi dal midollo allungato, a circa la prima vertebra lombare (non occupa quindi il canale vertebrale per tutta la sua lunghezza). Qui termina a forma di cono e prosegue verso il basso con un tratto connettivale privo di funzione (filo terminale).
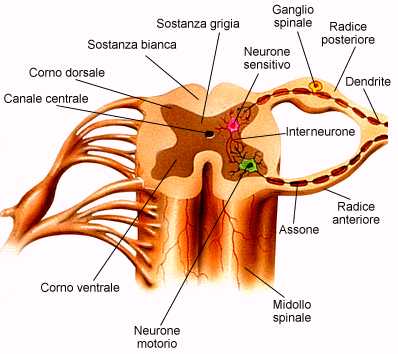
Al pari della colonna vertebrale, è diviso in segmenti (segmenti midollari). Sezionando trasversalmente uno di questi segmenti, si trova una disposizione oppposta a quella di cervello e cervelletto e uguale a quella del tronco encefalico. Infatti, nel midollo spinale la sostanza grigia è all'interno e la sostanza bianca all'esterno. La sostanza grigia del midollo spinale, composta soprattutto dai corpi delle cellule nervose (neuroni), ha una forma che ricorda una farfalla. Questi ammassi di corpi cellulari rappresentano i nuclei spinali. Nelle ali anteriori, dette corna anteriori, sono presenti, tra l'altro, i corpi cellulari dei motoneuroni alfa e gamma (classificazione delle fibre nervose), che innervano la muscolatura scheletrica. Gli assoni di questi neuroni fuoriescono dal midollo spinale, avvolti in una spessa guaina mielinica bianca, tramite le radici anteriori. A livello delle corna anteriori arrivano i segnali motori derivanti dalla corteccia cerebrale e dal tronco encefalico (nuclei vestibolari).
Le ali posteriori, corna posteriori, si estendono fino all'imbocco delle radici posteriori, dove si raccolgono, in un ganglio spinale, le fibre sensitive (sensibilità estero-propriocettiva). Qui terminano anche gli assoni deputati alla soppressione del dolore provenienti dal tronco encefalico. Secondo la teoria del "gate control" il neurotrasmettitore delle fibre del tatto, encefalina, inibisce la trasmissione sinaptica bloccando la percezione del dolore; stimoli emotivi intensi (shock) e stimoli tattili (digitopressione, agopuntura) possono così rendere temporaneamente insensibili al dolore.
I corni laterali sono piccole sporgenze a metà strada tra corna anteriore e posteriore. Le loro cellule, tramite le radici anteriori, inviano assoni agli organi innervati dal sistema nervoso vegetativo.
Al centro della sostanza grigia vi è un sottile canale centrale che è un residuo embrionale e non di rado è occluso o dilatato in cisti.
Immediatamente adiacenti alla sostanza grigia decorrono brevi fasci di fibre nervose, detti fasci fondamentali, che collegano 4-5 segmenti midollari confinanti circondando, come un sottile mantello, l'intera sostanza grigia. Essi provvedono a schemi di riflessi spinali complessi ovvero che non riguardano la muscolatura di un unico segmento (riflessi polisegmentali); tramite i riflessi spinali, ogni singolo segmento mdollare ha la capacità di controllare autonomamente funzioni motorie specifiche che richiedono rapidità di risposta.
La massa della sostanza bianca è invece formata da lunghi fasci midollari ascendenti e discendenti che collegano encefalo e midollo spinale. Questi fasci decorrono in tre cordoni a destra e tre a sinistra, separati fra loro dalla fessura centrale midollare, dal corno posteriore e da quello anteriore. L'ordine per una contrazione muscolare di precisione (ad esempio riguardanti movimenti delle dita della mano) viaggia lungo fibre che provengono dall'area motoria primaria encefalica attraverso il fascio piramidale del midollo spinale (fascio corticospinale) e lungo il fascio corticonucleare diretto ai nuclei dei nervi cranici del tronco encefalico. Il 90% delle fibre del fascio piramidale si incrocia nel midollo allungato, passando dal lato opposto (decussazione), e discende nella compagine del fascio corticospinale (piramidale) laterale. Il restante 10% non si incrocia nel midollo allungato e decorre nel fascio corticospinale (piramidale) anteriore, incrociandosi immediatamente prima dell' entrata nella sostanza grigia delle corna anteriori. Le fibre di entrambi i fasci terminano direttamente (meno di 1/3) oppure tramite un interneurone sulle cellule motorie delle corna anteriori, formando una sinapsi col motonerone alfa e attivando in tal modo un riflesso spinale.
Encefalo e midollo spinale formano il sistema nervoso centrale.
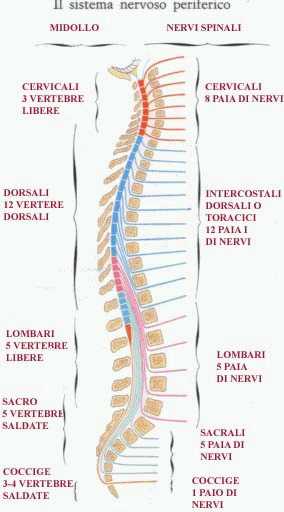
Ciascun segmento midollare emette, uno a destra e uno a sinistra, attraverso il foro di coniugazione o intervertebrale formato da due vertebre adiacenti, i nervi spinali che derivano ognuno dalla confluenza di una radice anteriore (fibre efferenti motorie con destinazione l'organo effettore, ghiandola o muscolo) e una posteriore (fibre afferenti sensitive, derivanti dai recettori sensoriali). Tutti i nervi spinali quindi risultano composti da una componente motoria e una sensitiva. A differenza della colonna vertebrale, il midollo spinale è dotato di 8 segmenti cervicali (invece di 7). I 12 segmenti toracici, 5 lombari, 5 sacrali e 1-2 coccigei sono invece in numero corrispondente alle relative vertebre. I nervi spinali cervicali fuoriescono dal foro di coniugacione superiore alla vertebra corrispondente, fatta eccezione dei due dell'VIII segmento (C8 dx e C8 sn) che escono da quello inferiore, così come accade per tutti i restanti segmenti. Affinchè ciò accada, essendo il midollo spinale più corto rispetto al canale vertebrale (a partire dal quarto mese di vita fetale il rachide si sviluppa più rapidamente del midollo spinale), le radici nervose dei segmenti lombari, sacrali e coccigei divengono sempre più distese verso il basso, formando così un fascio di sottili fibre nervose pressochè parallele che ricordano la coda di un cavallo (da cui la denominazione di cauda equina).
Appena fuori dal foro di coniugazione ogni nervo spinale si divide in 2 rami (entrambi sempre sia motori che sensitivi). Il ramo posteriore innerva, dal punto di vista motorio, la muscolatura spinale e, dal punto di vista sensitivo, la regione cutanea del dorso. Il ramo anteriore del nervo spinale costituisce la porzione più voluminosa e funzionalmente importante. Tutte le parti del corpo, a eccezione del viso, del tratto gastrointestinale e di parti della muscolatura cervicale (zone innervate dai nervi cranici), sono innervate dai nervi spinali. Insieme al sistema nervoso vegetativo, i nervi spinali formano il sistema nervoso periferico.
Il sistema nervoso vegetativo (autonomo, involontario) si divide in simpatico o ortosimpatico, parasimpatico e metasimpatico.
Il sistema nervoso ortosimpatico (simpatico) è composto da un'unità centrale di controllo, l'ipotalamo, e da una serie di coppie di neuroni efferenti (che emettono segnali in uscita). Il primo neurone (pregangliare), localizzato nel corno laterale dei segmenti spinali che vanno dalla prima vertebra toracica (T1) alla seconda lombare (L2), emette un assone che si diparte, attraverso la radice anteriore del midollo spinale, insieme al nervo spinale, da cui però si divide appena fuori dal foro di coniugazione spinale (foro intervertebrale) dirigendosi, come breve ramo comunicante bianco (fibre pregangliari mieliniche), al ganglio della catena dell'ortosimpatico del segmento spinale corrispondente (fa eccezione a questa regola l'innervazione della midollare del surrene). Le catene dell'ortosimpatico appaiono come due fili di perle poste parallelamente, una destra e l'altra a sinistra, lungo tutta la colonna vertertebrale. I suoi gangli (le "perle") sono connessi fra di loro da sottili fasci nervosi (ciò consente in realtà a una fibra pregangliare di prendere contatti anche con altri segmenti, sia superiori che inferiori). Da qui i segnali, direttamente o passando da ulteriori gangli, arrivano, tramite le fibre postgangliari (amieliniche), ai vari organi bersaglio. Le fibre pregangliari ortosimpatiche utilizzano come neurotrasmettitore l'acetilcolina e il peptide vasointestinale (VIP), quelle postgangliari la noradrenalina, l'adrenalina, la dopamina e il neuropeptide Y (NPY).
Anche il sistema nervoso parasimpatico è regolato dall'ipotalamo. Anatomicamente l'innervazione efferente parasimpatica decorre lungo i nervi cranici, di cui il X o nervo vago innerva gli organi della cavità toracica e addominale (quasi esclusivamente dipendenti da questo nervo) e i restanti raggiungono gli organi cranici, e lungo gli assoni dei neuroni parasimpatici presenti nei corni laterali dei segmenti spinali che vanno dalla seconda alla quarta vertebra sacrale (S2-S4). Questi ultimi decorrono poi quasi tutti nel nervo pudendo innervando organi genitali, vescica e intestino retto. Tutti i nervi parasimpatici contengono esclusivamente fibre pregangliari (che utilizzano come neurotrasmettitori l'acetilcolina e il peptide vasointestinale VIP) in quanto, a differenza del sistema nervoso ortosimpatico, i gangli parasimpatici sono situati in prossimità o all'interno degli organi innervati.
La maggior parte degli organi è innervata da entrambe le componenti del sistema nervoso vegetativo (ortosimpatica e parasimpatica). Vi sono però alcune parti del corpo che ricevono un’innervazione esclusivamente simpatica. Tra queste, la più importante è la porzione midollare delle surrenali (due piccole ma importantissime ghiandole endocrine poste ognuna sopra il polo superiore dei reni). La midollare del surrene viene inoltre innervata, attraverso i nervi splancnici, da fibre nervose pregangliari del simpatico che non si interrompono, ovvero che non formano sinapsi con i gangli della catena paravertebrale dell'ortosimpatico, ma che si connettono direttamente a un determinato gruppo di cellule della midollare, cellule cromaffini (formando in sostanza un ganglio interno), stimolandole a produrre le catecolamine adrenalina, noradrenalina e, in minima quantità, dopamina. Ci troviamo qui di fronte a una via direttissima che collega il cervello, tramite il sistema nervoso simpatico, alle surrenali. La visione del pericolo, infatti, mette in funzione questa via immediata (fase di allarme della reazione di stress), inducendo a produrre, da parte delle cellule della midollare del surrene, una miscela “allarmante” composta per l’80% di adrenalina e il 20% di noradrenalina, in quantità dieci volte maggiore del normale. Se però il pericolo perdura (fase di resistenza della reazione di stress), allora entreranno in scena ulteriori significativi cambiamenti (attivazione asse HPA) che coinvolgeranno ancora una volta le surrenali (questa volta la zona corticale), organi primari della reazione di stress.
Gli effetti del sistema nervoso ortosimpatico sono facilmente comprensibili se interpretati in relazione alla "reazione di attacco o difesa" (stress fisico e psichico). In molti casi il sistema nervoso parasimpatico è suo antagonista (schema reazione di stress).
Inoltre il sistema nervoso vegetativo invia al midollo spinale sensazioni quali, per esempio, dolore, stato di riempimento dello stomaco e della vescica. Tali fibre afferenti arrivano normalmente alla radice posteriore del midollo spinale non distinguendosi però in ortosimpatiche o parasimpatiche. Pertanto, poichè nei segmenti del midollo spinale convergono afferenze della sensibilità dolorifica degli organi interni e afferenze cutanee, il dolore viscerale puà essere proiettato e quindi percepito sulle aree cutanee, dermatomeri, del relativo segmento midollare (cuore-faccia interno braccio sinistro, cistifellea-addome superiore destro, pancreas-addome sinistro ecc.).
Descrivendo il sistema nervoso autonomo occorre ricordare l'esistenza di una terza componente vegetativa: il sistema nervoso metasimpatico, importante quanto le altre due (ortosimpatica e parasimpatica). Il sistema nervoso metasimpatico è presente all'interno delle pareti dell'intestino e dello stomaco come rete nervosa formata da circa cento milioni di neuroni, collegata alle fibre nervose ortosimpatiche e parasimpatiche (plesso sottomucoso di Meissner e motorio mienterico di Auerbach). Ricerche recenti hanno potuto affermare che questa rete svolge un ruolo in gran parte indipendente dal sistema nervoso centrale (SNC). Data l'importanza attruibuita dai fisiologi a tale struttura, essa viene definita come "cervello addominale o "cervello enterico". E' chiaro, data la grande attiguità, che l'alimentazione avrà sul sistema metasimpatico una grossa influenza.
In conclusione va detto che la classica netta separazione tra il sistema nervoso vegetativo, considerato come controllore delle funzioni viscerali in maniera indipendente dalla nostra volontà, e il sistema nervoso periferico volontario, è oggi sempre più messa in discussione da qualificati studiosi, in special modo dopo l'avvento della psiconeuroimmunologia. Il sistema nervoso costituisce una fittissima rete di comunicazione (solo nel cervello vi sono 10 miliardi di neuroni) diffusa in tutto il corpo e strettamente interconnessa con due altri importanti sistemi di regolazione dell'organismo: il sistema endocrino e il sistema immunitario.
Nel neurone la trasmissione del segnale lungo l'assone e il dendrite è di tipo elettrico (grazie a una variazione della polrarizzazione della membrana) mentre a livello sinaptico è di tipo chimico. I neurotrasmettitori (neuromediatori) sono molecole che consentono la trasmissione degli impulsi nervosi fra due neuroni attraverso le sinapsi o tra fibra nervosa terminale e organo effettore. Il neurotrasmettitore, all'arrivo dell'impulso nervoso (potenziale d'azione), viene liberato dalle vescicole presinaptiche, migra nello spazio intersinaptico fino a fissarsi su specifici recettori della membrana postsinaptica causando la depolarizzazione della membrana e quindi la trasmissione dell'impulso. Nel giro di pochi millisecondi viene disattivato, tramite specifici enzimi (colinesterasi, monoaminossidasi ecc.), e riassorbito nelle membrane presinaptiche (reuptake).
Esistono numerosi neurotrasmettitori con effetto eccitatorio e inibitorio. L'azione di uno specifico neurotrasmettitore dipende principalmente dalle caretteristiche del recettore postsinaptico (uno stesso neurotrasmettitore può avere effetti inibitori o eccitatori in base al recettore a cui si lega). Oggi sappiamo che un neurone è in grado di sintetizzare diversi neurotrasmettitori nelle diversi sinapsi che contrae, anche se utilizza sempre lo stesso nella medesima sinapsi.
I principali neurotrasmettitori sono: acetilcolina (presente nei neuroni di tipo colinergico quali i motoneuroni e neuroni effettori viscerali pregangliari), adrenalina, noradrenalina e dopamina (catecolamine secrete dalla midollare del surrene e dalle terminazioni nervose postgangliari del sistema nervoso orosimpatico), serotonina (con azione essenzialmente inibitoria, interviene, fra l'altro, nei ritimi sonno-veglia, nel controllo del dolore e delle funzioni ipotalamiche), aminoacidi GABA (acido-gamma-amino-butirrico, principale neurotrasmettitore inibitorio con presenza ubiquitaria), aspartato e glutammato (principali neurotrasmettitori eccitatori del sistema nervoso centrale). Oltre a questi vi sono molecole più complesse (vere e proprie proteine), i neuropeptidi, che possono fungere da neurotrasmettitori: glicina (neurotrasmettitore inibitorio, analogo al GABA, presente negli interneuroni del midollo spinale), peptide vasointestinale (VIP), neuropeptide Y (NPY), sostanza P, somatostatina, bombesina, istamina, neurotensina, encefalina e prostaglandina che, nel sistema nervoso centrale, possono fungere sia da neurotrasmettitori sia da neurormoni agendo a distanza su bersagli di natura endocrina. Talvolta i neuropeptidi possono agire da modulatori sinaptici (venendo secreti assieme ai neurotrasmettitori a livello sinaptico) organizzando i segnali specifici ed elementari dei normali neurotrasmettitori in un comportamento complesso.
I neurotrasmettitori hanno un ruolo essenziale, oltre che per la trasmissione degli impulsi nervosi, nella funzione di regolazione-modulazione dell'attività globale del sistema nervoso centrale, incluse le attività cognitive ed emotive.
In linea generale, la velocità di conduzione dell'impulso è tanto più elevata quanto maggiore è il diamentro della fibra nervosa ed esso va da 0,2 a 20 µm (1 µm = 1 micrometro, corrisponde a 1 milionesimo di metro), con velocità di conduzione da 0,5 a 120 metri/secondo. La vlocità di conduzione è inoltre aumentata in presenza della guaina mileinica.
Esistono due tipi di classificazione delle fibre nervose una classificazione generale, ossia che riguarda tutti i tipi di fibre, e una specifica per le fibre sensitive.
La classificazione generale (classificazione di Erlanger e Gasser) risulta:
Fibre di tipo o gruppo A, sono le tipiche fibre mieliniche dei nervi spinali e sono a loro volta suddivise in alfa, beta, gamma, delta, con diametro e quindi velocità di conduzione decrescente (velocità di conduzione da 120 a circa 6 m/s, diametro da 20 a 1 µm);
Fibre di tipo o gruppo B, sono le fibre pregangliari (mieliniche) del sistema nervoso autonomo, con valori di velocità di conduzione e diametro fra il tipo A e C.
Fibre di tipo o gruppo C, fibre amileiniche di piccolo diametro e quindi a bassa velocità di conduzione, che costituiscono più della metà delle fibre sensitive dei nervi periferici e la totalità delle fibre postgangliari del sistema nervoso autonomo (velocità di conduzione da 2 a 0,5 m/s, diametro inferiore a 1,2 µm).
La classificazione delle fibre sensitive (classificazione di Lloyd) è la seguente:
Fibre tipo Ia, provenienti dalle terminazioni anulospirali dei fusi neuromuscolari, corrispondono al tipo A alfa della classificazione generale (diametro medio 17 µm);
Fibre tipo o gruppo Ib, derivano dagli organi muscolo-tendinei del Golgi, corrispondono anch'esse al tipo A alfa (diametro medio 16 µm);
Fibre tipo o gruppo II, provenienti dalla maggior parte dei meccanorecettori per la sensibilità cutanea fine (esterocettori cutanei), e dalle terminazioni secondarie e a fiorame dei fusi neuromuscolari, corrispondono alle fibre A beta e gamma della classificazione generale (circa 8 µm di diametro);
Fibre tipo o gruppo III, responsabili della sensibilità tattile grossolana, della termica e delle sensazioni dolorifiche puntiformi, corrispondono alle fibre A gamma della classifica generale (circa 3 µm di diametro);
Fibre tipo o gruppo IV, amieliniche, trasportano impulsi della sensibilità dolorifica, termica, del prurito, tattile grossolana, appartengono al tipo C della classificazione generale (diametro 0,5-2 µm).
Tale differenzione delle fibre nervose è giustificata dal fatto che, in alcune situazioni, è indispensabile che determinati segnali nervosi giungano rapidamente al sistema nervoso centrale (come ad esempio durante la deambulazione o ancor più la corsa), in altri casi invece, come nell'informazione sensitiva relativa al dolore prolungato o cronico, sono sufficienti fibre nervose a conduzione molto lenta.
Il riflesso nervoso è una reazione del sistema nervoso centrale a stimolazioni esterne o interne (rilevate tramite i recettori sensoriali), atta a modificare lo stato di contrazione muscolare o di secrezione ghiandolare (reazione motoria, vasomotoria o ghiandolare). I riflessi interessano sia il sistema nervoso volontario (nervi spinali e cranici) che quello vegetativo (o autonomo). Il meccanismo di base, elementare, di un riflesso nervoso, definito arco riflesso o arco diastaltico, è costituito da: recettore sensoriale > via nervosa afferente o sensitiva > centro nervoso (posto nel sistema nervoso centrale) > via nervosa efferente o motoria > effettore (muscolo o ghiandola). Quando i riflessi nervosi hanno come centro nervoso il midollo spinale, si definiscono riflessi spinali, in altri casi i centri sono situati nel tronco encefalico (riflessi troncoencefalici).
I riflessi nervosi vengono suddivisi in semplici (detti anche incondizionati o congeniti) e complessi (o condizionati o acquisiti). Si individuano inoltre due grandi famiglie di riflessi: i riflessi propriocettivi, che originano dalla stimolazione dei propriocettori e che sono monosinaptici (riflesso di stiramento muscolare o riflesso miotatico o osteotendineo ROT, relativo ai fusi neuromuscolari) o dinaptici (riflesso miotatico inverso, relativo agli organi muscolo-tendinei del Golgi), a livello di un solo segmento midollare nel caso dei riflessi spinali, e i riflessi esterocettivi, che derivano dalla stimolazione degli esterocettori della cute e delle mucose, polisinaptici ossia, nel caso dei riflessi spinali, agenti a vari livelli midollari tramite i fasci fondamentali (riflesso plantare, corneale, faringeo, addominale, cremasterico, di retrazione ecc.).
Grazie ai riflessi, in particolare quelli semplici che presentano il grande vantaggio della rapidità (40 m/s per quelli rapidi), i livelli superiori possono demandare a midollo spinale e tronco encefalico, funzioni motorie specifiche che richiedono rapidità di risposta. Tuttavia, grazie alla modalità parallela, i centri encefalici superiori possono interagire direttamente sugli inferiori integrando e vicariando, in maniera immediata, funzioni; questo aspetto risulta fondamentale nel recupero funzionale di alcune lesioni del sistema nervoso centrale.
ll riflesso spinale avviene in genere grazie all'attività di specifici neuroni, collocati nel midollo spinale, detti interneuroni. Gli interneuroni si inseriscono, tramite sinapsi (connessione fra due neuroni), tra neuroni sensoriali (afferenti dai estero-propriocettori) e motori (efferenti). Gli interneuroni hanno perlopiù azione inibitoria (e talvolta facilitatoria) e sono di tre tipi (classificazione delle fibre nervose): (Ia) che coordinano l'attività di muscoli antagonisti (che esercitano azioni opposte), (Ib) correlati alle afferenze degli organi del Golgi che registrano la tensione tendinea e quindi la contrazione muscolare (da notare che l'influenza di questo tipo di interneurone dipende molto dalle afferenze cutanee a bassa soglia e articolari), cellule di Renshaw che hanno azione inibitoria diretta sul motoneurone. Durante gli atti motori, ogni muscolo che agisce su un'articolazione ha uno o più muscoli antagonisti che vengono contemporaneamente inibito (inibizione antagonista) o attivati (attivazione antagonista) tramite rispettivamente un interneurone inibitorio o facilitatorio interposto tra le fibre afferenti e i relativi motoneuroni alfa.
I riflessi spinali lavorano in modo gerarchicoo: controllo dei singoli muscoli, controllo della coordinazione muscolare riguardante una sola articolazione e riguardante più articolazioni. Mentre, tramite la modalità parallela, i centri superiori possono interagire direttamente sui riflessi integrandone e vicariandone, in maniera immediata, funzioni; ad esempio il midollo spinale da solo non è un grado di garantire una deambulazione fluida e sicura.
I riflessi svolgono un ruolo determinante in tutta la fisiologia dell'organismo e naturalmente anche nel sistema dell'equilibrio.
I riflessi spinali semplici vengono infine utilizzati in campo clinico per diagnosticare problematiche neurologiche.
Nessun commento:
Posta un commento